eISSN: 1989-3612
DOI: https://doi.org/10.14201/art2024.31935
NORMATIVIDAD NATURAL: EL “ES” Y EL “DEBE” DEL COMPORTAMIENTO ANIMAL1
TRADUCCIÓN
Frans B.M. DE WAAL
Living Links, Yerkes National Primate Research Center, Emory University, Atlanta, GA, United States
dewaal@emory.edu
RESUMEN: Suele suceder que la evolución del comportamiento se considera irrelevante para la comprensión de la moral humana, por el hecho de que carece de carácter normativo (el ‘debe’) y consiste enteramente en descripciones de cómo son las cosas o cómo sucedieron (el ‘es’). No obstante, el comportamiento que es producto de la evolución, incluido el de otros animales, no está completamente desprovisto de normatividad. Si se define la normatividad como la adhesión a un ideal o estándar, existe amplia evidencia de que los animales tratan a sus relaciones sociales de esta manera. En otras palabras, persiguen valores sociales. En este artículo reviso la evidencia sobre el hecho de que los primates no humanos intentan activamente preservar la armonía dentro de su red social, por ejemplo, reconciliándose después de un conflicto, protestando contra las divisiones desiguales y deteniendo peleas. Al hacerlo, corrigen las desviaciones respecto de un estado ideal. Sumado a ello, y con el fin de prevenir tales desviaciones, muestran autocontrol emocional y resolución anticipada de conflictos. El reconocimiento de la orientación hacia una meta y el carácter normativo del comportamiento social animal nos permite cerrar parcialmente la brecha entre el ‘es’ y el ‘debe’ erigida en relación con el comportamiento moral humano.
Palabras clave: moral, normatividad, resolución de conflictos, interés comunitario, justicia, aversión a la inequidad, control emocional.
1. INTRODUCCIÓN
Uno de los problemas más irritantes a los que se enfrentan los intentos de fundamentar la moral en la biología es la llamada división ‘ser/deber ser’. Si acaso se trata de una división real o no, depende en parte de cómo enunciemos la pregunta. Vale resaltar que quien la formulara inicialmente, David Hume (1739), no la vio, de hecho, como una división nítida. Hace ya casi tres siglos, lo que nos pidió fue que tuviéramos la precaución de no asumir que podemos derivar el ‘debe’ del ‘es’ y agregó que, si vamos a intentarlo, debemos dar una razón para ello. Habiendo notado la frecuencia con la que algunos autores pasaban de descripciones de cómo son las cosas a declaraciones sobre cómo deberían ser, agregó:
Este cambio es imperceptible, pero resulta, sin embargo, de la mayor importancia. En efecto, en cuanto que este ‘debe’ o “no debe” expresa alguna nueva relación o afirmación, es necesario que esta sea observada y explicada y que al mismo tiempo se dé razón de algo que parece completamente inconcebible, a saber: cómo es posible que esta nueva relación se deduzca de otras totalmente diferentes (Hume, 1739, 335).2
En otras palabras, la forma en que nos parece que deben comportarse los humanos no es simplemente un reflejo de la naturaleza humana. Así como no se pueden inferir las normas de tránsito de la descripción de un automóvil, tampoco se pueden inferir códigos morales a partir de saber quiénes o qué somos. El punto de Hume se entiende bien, pero está muy lejos de la ampliación que hicieron de él algunos filósofos posteriores, quienes convirtieron su pedido de cautela en la “guillotina de Hume”, proclamando un abismo infranqueable entre el ‘es’ y el ‘debe’ (Black, 1970). De ninguna manera existe acuerdo sobre este tema (sigue siendo un debate filosófico perenne), pero algunos han ido tan lejos empuñando esta guillotina que pretendieron acabar con todos y cada uno de los intentos, incluso los más cautelosos, de aplicar la lógica evolutiva o la neurociencia a la comprensión de la moral humana. La ciencia no puede decirnos cómo entender la moral, argumentan. Esto bien puede ser cierto, pero la ciencia sí puede ayudar a explicar por qué ciertos resultados son preferidos/favorecidos por sobre otros y, por lo tanto, por qué la moralidad es como es. Para empezar, no tendría sentido diseñar reglas morales que fueran imposibles de seguir, al igual que no tendría sentido hacer reglas de tránsito que los automovilistas no puedan obedecer. Esto se conoce como el argumento “el ‘debe’ implica el ‘puede’”. La moral debe amoldarse a la especie a la que está destinada.
El ‘es’ y el ‘debe’ son como el yin y el yang de la moralidad. Tenemos ambos, necesitamos ambos, no son lo mismo, pero no se pueden desvincular por completo. Se complementan entre sí (ver también Kitcher, 2014, en este número3). Al enfatizar la importancia de la naturaleza humana, el mismo Hume (1739) ignoró la ‘guillotina’ que lleva su nombre: vio la moral como un producto de las emociones, colocando la empatía (a la que llamó ‘simpatía’) en la parte superior de su lista. Esta opinión no representaba ninguna contradicción, ya que, como se mencionó más arriba, lo único que sugería era que tengamos prudencia en el paso del cómo somos al cómo debemos comportarnos (Baier, 1991). Nunca dijo que tal movimiento estuviera prohibido (aunque podría no haber estado de acuerdo con Singer [1973], para quien el debate sobre la división entre el ‘es’ y el ‘debe’ es una “trivialidad” que depende por completo de la definición de moral).
Si bien estoy de acuerdo con muchos filósofos en que es difícil, tal vez incluso imposible, razonar desde el nivel de cómo son las cosas hasta el cómo deberían ser, exploraré aquí si acaso la brecha divisoria resulta igualmente extensa cuando dejamos el dominio conceptual y entramos en el de las tendencias y motivaciones conductuales reales. ¿Qué tal si la moral no se construye racionalmente, sino que, como pensaba Hume, se basa en valores emocionales? ¿Qué tal si la biología no está sólo en el lado del ‘es’ de la ecuación, sino que también nos informa sobre el lado del ‘debe’, por ejemplo, al explicar qué valores perseguimos y por qué razón evolutiva? Cada organismo se esfuerza por lograr ciertos resultados. La supervivencia es uno, la reproducción es otro, pero muchos organismos también persiguen efectos sociales que se acercan a los respaldados por la moral humana.
El hecho de que el comportamiento animal no está libre de normatividad (definida como la adhesión a un ideal o estándar) apenas necesita argumentación. Tengamos en cuenta, por ejemplo, la reacción de la araña ante una red dañada. Si el daño es extenso, abandonará su telaraña, pero la mayoría de las veces entrará en modo de reparación: devolverá la telaraña a su estado funcional anterior, rellenando agujeros o ajustando los hilos dañados mediante otros nuevos (Eberhard, 1972). Del mismo modo, perturbar un nido de hormigas o un montículo de termitas conduce a una reparación inmediata, al igual que los daños generados a un dique de castores o a un nido de pájaros. La naturaleza está llena de estructuras físicas construidas por animales guiados por un patrón que indica cómo debería verse la estructura. Dicho patrón motiva la reparación o el ajuste tan pronto como la estructura se desvía del ideal. En otras palabras, los animales tratan estas estructuras de manera normativa. No estoy pensando necesariamente aquí en un juicio normativo. No está claro si los animales sienten la obligación de comportarse de una manera particular, ni asumo que cada individuo de una gran colonia tiene noción del nido como un todo, pero es innegable que los animales persiguen objetivos, colectiva o individualmente.
La cuestión aquí es si hacen lo mismo con respecto a las relaciones sociales y a la sociedad en general ¿Buscan ciertos resultados sociales y corrigen o desalientan las desviaciones de las expectativas? ¿Adoptan un enfoque normativo de las relaciones sociales y, de ser así, está guiado por el mismo tipo de emociones y valores que subyacen a la moral humana? Churchland (2011, 175) sugiere que hay una transición de las emociones sociales a los valores morales, sosteniendo que “las emociones básicas son la forma en que la Madre Naturaleza nos orienta para hacer lo que prudentemente debemos”. La pregunta es si este mismo movimiento es reconocible en otras especies.
2. JERARQUÍA SOCIAL Y CONTROL DE IMPULSOS
Lo opuesto a la moral es que simplemente hagamos “lo que queremos”, siendo un supuesto subyacente que lo que queremos no es lo moralmente bueno. Este sigue siendo un argumento religioso frecuente en contra de la ética naturalizada (Gallagher, 2004). Desde este punto de vista, la moral se basa en la capacidad exclusivamente humana de inhibir las tendencias naturales (Huxley, 1894). Por ejemplo, Kitcher (2006) etiquetó a los chimpancés como “caprichosos” [wantons], definiéndolos como criaturas vulnerables a cualquier impulso que incida sobre ellos. En algún momento de nuestra evolución superamos esta impulsividad caprichosa y es esa superación, justamente, la que nos hizo humanos. Según Kitcher (2006, 136), este proceso comenzó con la “consciencia de que ciertos tipos de comportamientos proyectados podrían tener resultados problemáticos”.
Sin embargo, innumerables animales viven con un tipo similar de conocimiento, no sólo cuando intentan evitar ser detectados por depredadores o presas mediante la supresión del sonido y el movimiento, sino también en el ámbito social. Una jerarquía de dominancia es un sistema gigante de inhibiciones sociales, que, sin duda, allanó el camino para la moral humana, el cual también es un sistema de este tipo. El control de los impulsos es clave para evitar “resultados problemáticos”. En macacos y otros primates, los machos de bajo rango varían su comportamiento dependiendo de la presencia o ausencia del macho alfa. Tan pronto como el alfa les da la espalda, los otros machos se acercan a las hembras. En un experimento que ponía a prueba este principio, los machos de bajo rango se negaban a acercarse a las hembras mientras el dominante miraba desde el interior de una caja transparente, pero tan pronto como este macho era retirado, copulaban libremente con ellas. Los machos de bajo rango también aprovechaban la ocasión para realizar las típicas demostraciones de vigor [bouncing displays] de los machos de alto estatus. Después de tales episodios, sin embargo, estaban excesivamente nerviosos al reunirse con el macho alfa, mostrando los dientes con tanta sumisión al saludarlo que los experimentadores interpretaron su comportamiento como un reconocimiento implícito de que habían violado un código social (Coe y Rosemblum, 1984). Tal vez las reglas sociales no se obedezcan simplemente en presencia de los dominantes y se olviden en su ausencia, sino que hasta cierto punto también se interioricen. Ahora bien, cuando los científicos han tratado de medir el grado de internalización de las reglas impuestas por humanos en los perros, estudiando su comportamiento de aspecto culpable después de las violaciones a las mismas, no han encontrado mucho más que cierto efecto directo del comportamiento del dueño sobre el perro (Vollmer, 1977; Horowitz, 2009).
No sólo los individuos de bajo rango, sino también los de alto rango, se benefician del control de impulsos. Por ejemplo, un chimpancé macho alfa experimentado (Pan troglodytes) puede ser desafiado de modo evidente por parte de un macho más joven, que arroja piedras en su dirección o hace una llamativa exhibición de fuerza con todos los pelos de punta (este es un modo de poner a prueba los nervios del alfa) y, sin embargo, ignorar totalmente el estruendo, como si apenas se diera cuenta, lo que obliga a su retador o a rendirse o a intensificar aún más la situación (de Waal, 1982).
En última instancia, las inhibiciones asociadas con la jerarquía llegan a través del castigo. Por ejemplo, después de haber privado de agua a una gran manada de monos rhesus (Macaca mulatta) durante tres horas, se puso a su disposición un único estanque lleno de agua. Todos los adultos fueron a beber en orden jerárquico. No obstante, los infantes y los jóvenes bebían con los machos de más alto rango y se mezclaban con la línea matricial superior, ignorando así la jerarquía social. Es recién en el tercer año de vida, a través de exclusiones y castigos cada vez mayores, que los menores comienzan a aprender su lugar en el orden de rango general y a converger así con el rango de su madre (de Waal, 1993).
En el caso de los simios, dado que se desarrollan más lentamente que los monos, los jóvenes andan prácticamente impunes durante sus primeros cuatro años de vida. Por ejemplo, usan la espalda de un macho dominante como trampolín, roban comida de las manos de otros o golpean a un joven mayor tan fuerte como pueden: nada de lo que hacen resulta/parece incorrecto. Uno puede imaginar la conmoción de un joven cuando es rechazado o castigado por primera vez. Los castigos más dramáticos son los de aquellos varones jóvenes que se aventuran demasiado cerca de una hembra sexualmente atractiva (de Waal, 1982; Figura 1). Los machos jóvenes sólo necesitan una o dos lecciones de este tipo. A partir de allí, todo macho adulto puede hacer que salten lejos de una hembra simplemente con una mirada o un paso adelante. De esta manera, los más jóvenes aprenden a controlar sus impulsos sexuales o, por lo menos, se vuelven más discretos a la hora de actuar cuando son guiados por ellos.

Figura 1. Un macho joven, de unos 4 años, ha mostrado demasiado interés por una de las hembras en celo y ahora está siendo castigado por un macho adulto, que le ha metido el pie en la boca y lo balancea. Esto servirá como una lección para el resto de la vida del joven macho sobre la competitividad de los machos en torno a las hembras sexualmente atractivas. Fotografía de Frans de Waal
La capacidad para controlar los impulsos puede testearse experimentalmente de la misma manera que se testea la demora de la gratificación en niños (Mischel et al., 1972; Logue, 1988). A los niños se les da un malvavisco con la promesa de que, si no se lo comen, tendrán otro más. Muchos niños tienen la capacidad de esperar durante minutos. De manera similar, tanto los simios (Beran et al., 1999) como los monos (Amici et al., 2008) dejan pasar una recompensa inmediata en favor de una mejor y más tardía. También se ha mostrado que los chimpancés, como los niños, juegan más con sus juguetes en presencia de recompensas que se acumulan, lo que sugiere que intentan auto-distraerse frente a la tentación. Esta acción permite a los simios demorar la gratificación hasta 18 minutos (Evans y Beran, 2007). Otros estudios han demostrado que los simios pueden anular un impulso inmediato en favor de necesidades futuras, un aspecto esencial para una planificación exitosa de la acción (Osvath y Osvath, 2008).
El mismo entrelazamiento entre emoción y cognición reconocido en humanos parece estar presente en nuestros parientes cercanos, incluyendo el control deliberado de las emociones. En la medida en que dicho control está mediado por los lóbulos frontales, es importante señalar que la opinión popular de que esta parte del cerebro está excepcionalmente desarrollada en nuestra especie es errónea. El cerebro humano es esencialmente un cerebro de mono ampliado linealmente (Herculano-Houzel, 2009; Barton y Venditti, 2013).
3. NORMATIVIDAD UNO A UNO
La moral puede ser definida como un sistema de reglas que gira en torno a ayudar o, al menos, a no dañar a los demás seres humanos. Su propósito es el bienestar de los demás y, a menudo, antepone los intereses de la comunidad frente a los del individuo. No niega el interés propio, pero pone restricciones a su búsqueda con el fin de promover una sociedad cooperativa (de Waal, 1996, 2006). Esta definición funcional diferencia a la moral de las costumbres y los hábitos, como comer con cuchillo y tenedor versus comer con palillos o con las manos desnudas. La distinción entre reglas morales y convenciones ya se ve claramente en niños pequeños (Killen y Rizzo, 2014, este número). En otro trabajo he distinguido dos niveles de reglas morales: (a) reglas en el nivel uno a uno (diádicas) de las relaciones sociales, y (b) reglas en el nivel comunitario (de Waal, 2013). La Tabla 1 resume ejemplos en el nivel uno a uno.
Ideal |
Desviación |
Reparatión o correctión |
Restauración |
Jerarquía |
Desobediencia o desafío al rango |
Castigo o restablecimiento de dominancia |
Annonía |
Relatión cercana |
Conflicto |
Reconciliatión |
Armonía |
Cooperatión |
Recompensa desigual |
Protesta o compartición |
Annonía |
Juego relajado |
Compañero lastimado |
Señales de remediación |
Annonía |
Tabla 1. Cuando los individuos buscan preservar relaciones sociales armoniosas, aplican la normatividad uno a uno. Su comportamiento refleja el valor que atribuyen a las buenas relaciones. Esta tabla ofrece cuatro ejemplos: restauración de la jerarquía de dominancia, reparación de relaciones, reacciones negativas a la inequidad y reanudación del juego. En todos los casos, los primates y otros animales buscan activamente que una relación social retorne a su estado original
3.1. Reconciliación
El nivel uno a uno gira en torno a la preservación de relaciones valiosas. Una de sus expresiones más comunes es la resolución de conflictos, reportada por primera vez por de Waal y van Roosmalen (1979). Un ejemplo típico refiere a dos chimpancés machos que luego de haber estado persiguiéndose y gritándose, descansan en un árbol (Figura 2). Diez minutos después, un macho extiende su mano al otro, rogándole un abrazo. En cuestión de segundos, se abrazan, se besan y bajan juntos al suelo para acicalarse. Entendiéndolo como una reconciliación, este proceso se define como un contacto amistoso que ocurre poco después de un conflicto entre dos partes. Mientras que el besarse es la forma más típica de reconciliación entre chimpancés, los bonobos lo hacen mediante el comportamiento sexual (de Waal, 1987) y los macacos rabones llevan a cabo un comportamiento ritual en el cual esperan hasta que el subordinado se presente para luego sostener sus caderas (de Waal y Ren, 1988). Cada especie tiene su propio camino, pero el principio básico sigue siendo el mismo: los oponentes vuelven a reunirse después de una pelea.

Figura 2. La situación después de un prolongado y ruidoso conflicto entre dos machos adultos en un zoológico. El macho desafiado (izquierda) había huido hacia el árbol. 10 minutos después su oponente le extendió una mano. En cuestión de segundos, los dos machos mantuvieron un reencuentro físico. Fotografía de Frans de Waal
La primatología se ha interesado durante mucho tiempo en las relaciones sociales, por lo que la idea de reparación de relaciones, implícita en la etiqueta de reconciliación, atrajo rápidamente su atención. Ahora sabemos que una treintena de especies diferentes de primates se reconcilian después de las peleas y que esa reconciliación no se limita a los primates. Hay pruebas de este mecanismo en hienas, delfines, lobos, cabras domésticas, etc. La razón por la que la reconciliación está tan extendida es que restituye relaciones que han sido dañadas por la agresión pero que, sin embargo, son esenciales para la supervivencia. Dado que muchos animales establecen relaciones de cooperación en las que ocasionalmente surgen conflictos, los mecanismos de reparación son esenciales. El creciente campo de la resolución de conflictos en animales ha sido examinado por de Waal (2000) y Aureli y de Waal (2000).
La mayoría de estos estudios apoyan la Hipótesis de la Relación Valiosa, que puede formularse de la siguiente manera: “La reconciliación ocurrirá especialmente entre individuos que tienen mucho que perder si su relación se deteriora”. Esta hipótesis también ha sido respaldada por un elegante experimento que manipuló el valor de la relación, promoviendo la cooperación entre monos y aumentando de este modo su disposición a reconciliarse después de las peleas (Cords y Thurnheer, 1993). Las ideas anteriores se han formalizado en el Modelo Relacional, que sitúa el conflicto en un contexto social. La agresión es vista como una de las varias opciones para resolver un conflicto de intereses. Otras opciones son evitar al adversario (común en especies jerárquicas y territoriales) y compartir recursos (común en especies tolerantes). Sopesando los costos y beneficios de cada opción, el conflicto puede escalar hasta el punto de la agresión, después de lo cual todavía existe la opción de reparar el daño por medio de la reconciliación, la cual suele verse favorecida cuando las partes tienen intereses superpuestos (de Waal, 2000; Figura 3). Aplicando la misma metodología primatológica estandarizada a niños humanos, se han obtenido resultados muy similares (Verbeek et al., 2000).
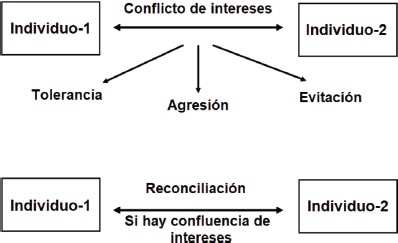
Figura 3. De acuerdo con el modelo relacional, el comportamiento agresivo es una de las varias formas en que se pueden resolver los conflictos de intereses. Otras formas posibles son la tolerancia (p. ej., compartir recursos) y evitar la confrontación (p. ej., por parte de los subordinados a los dominantes). Si ocurre una agresión, dependerá de la naturaleza de la relación social el que se lleven a cabo o no intentos de reparación. En el caso de que exista un fuerte interés mutuo en mantener la relación, es probable que haya una reconciliación. Las partes negociarán los términos de su relación pasando por ciclos de conflicto y reconciliación. Véase de Waal (2000)
3.2. Resolución preventiva de conflictos
El hecho de que los primates se protegen contra los efectos perjudiciales del conflicto y la angustia es algo que resulta visible durante el juego. Cuando los jóvenes tienen edades muy diferentes, los juegos a menudo se vuelven demasiado brutos para el compañero más joven, como cuando tuercen sus piernas o cuando un mordisqueo se convierte en un tarascón. A la menor señal de peligro, su madre interrumpirá el juego. Normalmente, excepto por las roncas y jadeantes risas de los simios (semejantes a la risa humana), el juego es completamente silencioso (van Hooff, 1972). Tras registrar cientos de estas escenas de lucha descubrimos que los chimpancés jóvenes emiten este tipo de vocalización especialmente cuando la madre de un compañero de juegos más joven está mirando. Es decir, lo hacen más en presencia de la madre que cuando están solamente con el chimpancé más joven. Puede que las vocalizaciones en cuestión tengan la función de evitar la intervención materna, tranquilizándola sobre la naturaleza benigna de la interacción (Flack et al., 2004).
Por su parte, Bekoff (2001) analizó videos de juegos entre perros, lobos y coyotes, concluyendo que el juego de los cánidos está sujeto a reglas, genera confianza, requiere considerar al otro y enseña a los jóvenes a comportarse. El altamente estereotipado comportamiento de “reverencia de juego” (un animal se agacha sobre sus extremidades anteriores mientras levanta su trasero), ayuda a diferenciar el juego del sexo o el conflicto, con los cuales puede llegar a confundirse. El juego cesa abruptamente, sin embargo, tan pronto como un compañero hiere a otro. El transgresor precisará realizar una nueva reverencia, después de lo cual el compañero podría continuar el juego. Bekoff establece un paralelo con la moral:
Durante el juego social, mientras los individuos se divierten en un entorno relativamente seguro, aprenden reglas básicas que resultan aceptables para los demás: qué tan fuerte pueden morder, qué tan bruscamente pueden interactuar y cómo resolver conflictos. Es muy importante jugar de manera justa y confiar en que los demás también lo harán. Hay códigos de conducta social que regulan lo que está permitido y lo que no, y la existencia de estos códigos podría tener algo que decir sobre la evolución de la moral (Bekoff, 2001, 85).
El comportamiento destinado a la preservación de las buenas relaciones es un indicio del gran valor que se le otorga a la armonía social. Kummer (1995) ofrece observaciones sorprendentes de la forma en que los líderes de harén de los babuinos hamadryas (Papio hamadryas), al encontrarse frente a un árbol frutal demasiado pequeño como para alimentar a sus dos familias, suspenderán su inevitable confrontación huyendo literalmente el uno del otro, seguidos por sus respectivas hembras y crías. Los machos de chimpancé enfrentan un dilema similar. Varios de ellos pueden sentarse cerca de una hembra que anuncia su fertilidad con los genitales hinchados. En lugar de competir, los machos mantienen activamente la paz. Aunque con frecuencia echan un vistazo a la hembra, pasan el día acicalándose mutuamente. Recién cuando todos están lo suficientemente relajados, uno de ellos intentará aparearse (de Waal, 1982).
Si bien las descripciones anteriores son cualitativas, también se han cuantificado las técnicas de prevención de conflictos. Tras una sugerencia inicial de de Waal (1987), según la cual habría un pico de acicalamiento entre los bonobos en cautiverio justo antes de la hora de comer, precediendo la posible competencia y tensión, los estudios han procurado medir el comportamiento en torno a la llegada de alimentos, que en la mayoría de los zoológicos e instituciones ocurre en un momento predecible del día. Los chimpancés se acicalan más mientras esperan comida y participan en “celebraciones” marcadas por altos niveles de contacto corporal tranquilizador una vez que llega la comida (de Waal, 1992a; Koyama y Dunbar, 1996). Los bonobos, por otro lado, muestran un aumento en su comportamiento de juego antes de la comida y una gran cantidad de contacto sociosexual a su llegada (de Waal, 1987; Palagi et al., 2006). Por lo tanto, los primates pueden anticipar la competencia por la comida y trabajan activamente para reducirla.
3.3. Persiguiendo una división justa de recompensas
Las reacciones negativas a las distribuciones de recompensas sesgadas, también conocidas como aversión a la inequidad (AI), son otra parte del mantenimiento de las relaciones diádicas. Los animales cooperativos precisan vigilar qué beneficios obtienen en relación con sus socios cooperativos, a fin de evitar que se aprovechen de ellos. En ausencia de una distribución equitativa, la cooperación mutualista podría convertirse fácilmente en una forma de altruismo por parte de quienes ganan menos. Este ‘problema de resultado’ [outcome problem] ha sido reconocido en humanos (Fehr y Schmidt, 1999), y es cada vez más un tema recurrente en la investigación con animales (Brosnan, 2011).
Los monos capuchinos son tan sensibles a la inequidad que ante las recompensas que se encuentran amontonadas espacialmente (resultando por ello monopolizables por las partes dominantes) se reducen más sus tendencias cooperativas que en el caso de recompensas dispersas (de Waal y Davis, 2003). Brosnan y de Waal (2003) probaron la AI en un experimento simple en el que dos monos recibían recompensas iguales o desiguales por la misma tarea, por ejemplo, uno recibía rodajas de pepino y el otro uvas (preferidas por ellos). Los autores encontraron que, cuando ambos recibían lo mismo, los individuos que obtenían la recompensa menor no se veían afectados, pero si su compañero recibía un mejor trato, a menudo se negaban a realizar o aceptar la recompensa. Se encontraron resultados similares en chimpancés (Brosnan et al., 2005). Sin embargo, las replicaciones experimentales que no requerían que los participantes desempeñasen una tarea, no dieron los mismos resultados (Braüer et al., 2006; Roma et al., 2006) (incluso en el caso de un estudio con los mismos monos que en el estudio original) (Dindo y de Waal, 2006). Por lo tanto, como podría predecirse desde una explicación evolutiva centrada en el desempeño de tareas y en la cooperación, las recompensas desiguales provocan reacciones negativas sólo en el contexto de una tarea que requiere esfuerzo. Finalmente, van Wolkenten et al. (2007) demostraron que las respuestas a la inequidad son realmente sociales en el sentido de que no pueden explicarse como reacciones negativas a recompensas menores cuando otras mejores están visibles. Es decir, la mera visibilidad tuvo poco efecto: las reacciones negativas ocurrieron sólo si las mejores recompensas eran consumidas por un compañero.
Se han observado respuestas AI similares en otras especies, tanto primates como no primates (Brosnan, 2011; Price y Brosnan, 2012; Range et al., 2012). Sin embargo, existe una restricción: hasta ahora, la mayoría de los estudios sólo se refieren a la AI por parte del individuo que recibe menos, conocida como AI desventajosa. En la AI ventajosa, en cambio, los sujetos responden negativamente al recibir una recompensa mayor que la de su compañero. Los humanos muestran tanto la última respuesta como la primera. Brosnan y de Waal (2012) especulan que la AI ventajosa, que indica un pleno sentido de justicia, ocurre cuando los individuos anticipan las implicaciones negativas de la AI desventajosa en los demás. Con el fin de proteger la relación frente a los efectos erosivos de las tensiones surgidas cuando un individuo recibe menos que el otro, el que recibe más intenta evitarlo igualando la recompensa. Los autores señalan esto como un sentido de justicia de segundo orden: “Para prevenir conflictos al interior de relaciones cercanas o beneficiosas, el individuo aventajado se beneficiará de protestar o rectificar la situación” (Brosnan y de Waal, 2012, 341).
Hasta el momento no hay signos de justicia de segundo orden en monos, como por ejemplo en los monos capuchinos del estudio original. En los simios, sin embargo, la evidencia va en aumento. La primera señal provino de un estudio de Brosnan et al. (2010) en chimpancés, en el que no sólo se negaban regularmente a realizar o aceptar sus recompensas los participantes que recibían la menor recompensa, sino también respondían de este modo quienes que recibían la mejor recompensa. En otras palabras, cualquier inequidad, no sólo la desventajosa, era resistida. Tenía sentido, por tanto, testear a los chimpancés con el Juego del Ultimátum (JU), que es el estándar de oro del sentido de justicia humano. En el JU, un individuo (el Proponente) puede dividir el dinero con otro individuo (el Respondedor). Si el Respondedor acepta la oferta, ambos jugadores son recompensados con la división acordada. Sin embargo, si el Respondedor rechaza la oferta, ninguno de los jugadores es recompensado. En las culturas occidentales, las personas suelen ofrecer alrededor del 50% de la cantidad disponible (Guth, 1995; Camerer y Lowenstein, 2004), al igual que en la mayoría de las otras culturas (Henrich et al., 2001). En contraste, un estudio del JU en chimpancés encontró que comparten la menor cantidad posible con los demás (Jensen et al., 2006). Sin embargo, la metodología de este experimento se desvió sustancialmente del JU humano típico y no estaba claro si los simios entendieron completamente la tarea.
Para superar estas objeciones, Proctor et al. (2013) diseñaron un procedimiento del JU que resultaba más intuitivo tanto para chimpancés como para niños humanos de 3 a 5 años. A los Proponentes se les presentó la posibilidad de elegir entre dos fichas de diferente color que podrían intercambiar por comida con un experimentador humano. Mientras que un color representaba una distribución equitativa de la recompensa (3 rodajas de plátano versus otras 3), el otro representaba una distribución desigual a favor del Proponente (5 rodajas de plátano versus 1). El Proponente tenía que entregar la ficha a su socio, el Respondedor, sentado detrás de una reja de malla. Quienes respondían podían aceptar la ficha y devolvérsela al experimentador, o rechazarla no devolviendo la ficha. Como en el JU humano típico, los Proponentes necesitaban la colaboración del Respondedor.
Las elecciones de fichas se compararon con elecciones en presencia de Respondedores que carecían de cualquier influencia. Los chimpancés resultaron sensibles a las contingencias del juego, de la misma forma que los humanos. Si su compañero tenía el control, dividían las recompensas en partes iguales con mayor frecuencia. Sin embargo, en ausencia de la influencia de su compañero, preferían la opción que les otorgaba la mayor proporción de recompensa. Siendo que los niños se comportan de manera similar, el estudio sugiere que los humanos y los chimpancés comparten patrones de toma de decisiones proactivas en relación con los resultados justos (Proctor et al., 2013).
4. PREOCUPACIÓN COMUNITARIA
En comparación con lo que ocurre en el caso de la normatividad individual uno a uno, hay muchos menos signos de normatividad a nivel comunitario en primates no humanos. Este es el nivel en que la moral humana parece ser única, ya que habitualmente extendemos nuestro razonamiento moral hacia la sociedad en su conjunto, especulando qué le sucedería a nuestra comunidad si acaso todos actuáramos de cierta manera. De hecho, solemos extender nuestro sistema de valores a interacciones en las que no estamos involucrados de manera directa. Siguiendo a Westermarck (1917, 238), las emociones morales difieren de las ordinarias “por su desinterés, aparente imparcialidad y generalidad”. Mientras que las emociones típicas conciernen sólo a nuestros intereses personales —cómo nos han tratado o cómo queremos que nos traten—, las emociones morales van más allá. Se ocupan del bien y del mal en un nivel más abstracto. Sólo cuando realizamos juicios respecto de cómo debería ser tratado alguien bajo ciertas circunstancias, estamos hablando de un juicio moral. Para transmitir la misma idea, Smith (1759) nos pidió que imagináramos cómo un “espectador imparcial” juzgaría el comportamiento humano.
Esto no quiere decir que el nivel de la normatividad comunitaria esté completamente ausente del comportamiento de nuestros parientes cercanos. En un trabajo anterior he denominado este nivel como el de la “preocupación comunitaria” (de Waal, 1996). Existen muchos ejemplos de vigilancia y mediación imparciales que parecen reflejar valores a nivel de la comunidad. En algunas especies, las intervenciones de los miembros de rango más alto ponen fin a las peleas o al menos reducen la gravedad de la agresión. A menudo, los chimpancés machos de alto rango juegan este papel en peleas entre hembras y/o entre jóvenes de su grupo (de Waal, 1982). Por ejemplo, si dos jóvenes están jugando y se desata una pelea, el macho alfa puede acercarse al área del conflicto para detener la pelea. Al suspenderla antes de que las madres intervengan y comiencen a pelear entre ellas, reduce los niveles de agresión dentro del grupo y evita también que la pelea de los jóvenes se intensifique.
Este patrón de comportamiento se conoce como “rol de control” (cf. Bernstein y Sharpe, 1966). De Waal (1982) proporcionó descripciones y análisis detallados, junto con datos que muestran que, durante la adopción de dicho rol, los machos ignoran sus lazos sociales con los participantes del conflicto. Mientras que la mayoría de los individuos apoyan a sus amigos y parientes, los machos controladores intervienen independientemente de sus preferencias sociales habituales (de Waal, 1992b). La capacidad de dejar de lado tales preferencias sugiere una forma rudimentaria de justicia en los sistemas sociales de los primates no humanos. También se ha observado vigilancia imparcial en chimpancés salvajes (Boehm, 1994), y un estudio reciente que comparó este comportamiento en varios grupos cautivos concluyó que estabiliza la dinámica social (von Rohr et al., 2012). Un estudio experimental de Flack et al. (2005), en el que se separó temporalmente del resto del grupo a miembros clave en lo que al control se refiere, mostró su importancia para el mantenimiento del aseo personal, el juego y otros signos de una sociedad armoniosa.
Otro método importante de resolución de conflictos que se ha identificado en grupos de primates es la mediación. La mediación ocurre cuando, en medio de un conflicto, un tercero se convierte en el puente entre dos oponentes incapaces de reconciliarse sin ayuda externa. La misma es caracterizada en el siguiente ejemplo (de Waal y van Roosmalen, 1979):
A veces, especialmente después de conflictos graves entre dos machos adultos, los dos oponentes eran reunidos por una hembra adulta. La hembra se acercaba a uno de los machos, lo besaba, lo tocaba o se presentaba ante él y luego, si el macho en cuestión la seguía, caminaba lentamente hacia el otro macho. El primero la seguía muy de cerca (a menudo inspeccionando sus genitales) y sin mirar al otro. En algunas ocasiones, la hembra miraba hacia atrás a su seguidor y, a veces, volvía hacia el que se había quedado atrás tirando de su brazo para hacer que la siguiera. Luego de que la hembra se sentaba cerca del otro macho, ambos comenzaban a acicalarla y simplemente continuaban acicalándose entre sí cuando ella se iba.
Llamar a tal comportamiento una expresión de interés comunitario de ninguna manera implica que no haya beneficios para el ejecutante. En los animales que viven en sociedad existe una gran superposición entre intereses comunitarios e individuales, de allí que cada individuo seguramente tenga interés en que su comunidad alcance cierto nivel de armonía y cooperación. Por lo tanto, el término ‘preocupación comunitaria’ no implica ningún sacrificio y, menos aún, una selección a nivel grupal. Simplemente establece que los individuos pueden promover los intereses de su comunidad como un todo, lo que bien puede ser para su propio beneficio al mismo tiempo que beneficia a otros.
Finalmente, el prestigio y la reputación son una parte fundamental del por qué los seres humanos a menudo actúan en nombre de la comunidad, incluso cuando no se benefician directamente de ello. En los simios pueden verse algunos destellos de reputación. Por ejemplo, si una pelea importante se sale de control, los espectadores van a despertar al macho alfa dándole algunos golpecitos. Conocido como el árbitro más eficaz, se le insta a intervenir. También prestan atención a cómo un individuo trata a otro, como sucede en los experimentos en los que prefieren interactuar con un humano que ha mostrado una actitud positiva hacia los demás, por ejemplo, el haber compartido comida con otros simios (Russell et al., 2008; Subiaul et al., 2008; Herrmann et al., 2013). En nuestros estudios, descubrimos lo siguiente: cuando dejábamos a la colonia observar a dos chimpancés que utilizaban trucos diferentes pero igualmente simples para obtener recompensas, preferían seguir al modelo de estatus superior. Este comportamiento evidenciaba el denominado ‘efecto de prestigio’, según el cual los simios imitan preferentemente a los miembros destacados de su comunidad (Horner et al., 2010).
Esta evidencia sugiere que los chimpancés realizan acciones que benefician a la comunidad en su conjunto y, a su vez, que poseen reputaciones individuales relacionadas con cómo ciertos individuos tratan a los demás o cuán dignos de imitación son. No obstante, dichos comportamientos están todavía muy lejos del interés humano por los estándares de la comunidad y por el bienestar del conjunto. Es especialmente en el nivel del interés comunitario y de la construcción de la reputación que los sistemas morales humanos se desvían de la normatividad hallada en otros primates.
5. CONCLUSIÓN
A la luz de lo desarrollado y en relación con la división entre el ‘es’ y el ‘debe’, la postura que supone que la biología, incluido el comportamiento animal, reside completamente en el lado del ‘es’, resulta difícil de mantener. Obviamente, podemos describir el comportamiento animal dejando de lado todas y cada una de las referencias a objetivos, intenciones y valores (del mismo modo que podemos describir el comportamiento humano de esta manera), pero tales descripciones pasan por alto un aspecto básico: los primates no humanos, así como también muchos otros animales, persiguen resultados específicos, tanto en relación con la construcción de ciertas estructuras físicas, por ejemplo nidos y redes, como en relación con los vínculos sociales. Intentan activamente preservar la armonía dentro de su comunidad social y con frecuencia corrigen las desviaciones de este ideal, por ejemplo, reconciliándose después de un conflicto, protestando contra divisiones desiguales o deteniendo peleas. Al corregir o tratar de corregir las desviaciones de un estado ideal, se comportan normativamente. A su vez, con el fin de prevenir dichas desviaciones, muestran autocontrol emocional y resolución anticipada de conflictos. Esto conlleva que el paso del comportamiento de los primates a las normas morales humanas sea un salto menor de lo que comúnmente se supuso.
Sin embargo, es probable que siga habiendo diferencias. Los otros primates parecen no extender las normas más allá de su entorno social inmediato y parecen no preocuparse por las relaciones sociales o situaciones en las que no participan de manera directa. A diferencia de los humanos, es posible que no sientan ninguna obligación de ser buenos ni que experimenten culpa y vergüenza cuando se equivocan. No sabemos si otros animales experimentan tales sentimientos vinculados con el ‘debe’. Es posible argumentar que, si bien su comportamiento podría entenderse como normativo en el sentido de que busca ciertos resultados, lograrían llevarlos a cabo sin la necesidad de realizar un juicio normativo. Es decir, podrían evaluar el comportamiento social como exitoso o no en relación con la consecución de sus objetivos, pero no en términos de lo correcto y lo incorrecto. Por otro lado, su comportamiento a veces sugiere una suerte de evaluación de acciones pasadas, como cuando un bonobo muerde a otro y poco después se le acerca, recordando el lugar exacto de la mordedura, sólo para pasar media hora lamiéndole la herida infligida (de Waal, 1989). Sin embargo, dada la inaccesibilidad de la experiencia animal, la existencia de una normatividad internalizada sigue siendo altamente especulativa. Por el momento, este artículo hace una afirmación más débil: en la medida en que el ‘debe’ de la moral humana refleja una preferencia por ciertos resultados sociales sobre otros, existen preferencias similares que parecen guiar a otros animales sin que ello implique necesariamente que a ellas subyazca el mismo sentido de obligación que guía a los seres humanos.
REFERENCIAS BIBLIOGRÁFICAS
Amici, F., Aureli, F. y Call, J. (2008). Fission-fusion dynamics, behavioral flexibility, and inhibitory control in primates. Curr. Biol., 18, 1415-1419. https://doi.org/10.1016/j.cub.2008.08.020
Aureli, F. y de Waal, F. B. M. (2000). Natural conflict resolution. Berkeley, CA: University of California Press. https://doi.org/10.1525/9780520924932-003
Baier, A. C. (1991). A progress of sentiments: reflections on Hume’s treatise. Cambridge, MA: Harvard University Press. https://doi.org/10.4159/9780674020382
Barton, R. A. y Venditti, C. (2013). Human frontal lobes are not relatively large. Proc. Natl. Acad. Sci. USA, 110, 9001-9006. https://doi.org/10.1073/pnas.1215723110
Bekoff, M. (2001). Social play behaviour cooperation, fairness, trust, and the evolution of morality. J. Consc. Studies, 8, 81-90.
Beran, M. J., Savage-Rumbaugh, E. S., Pate, J. L. y Rumbaugh, D. M. (1999). Delay of gratification in chimpanzees (Pan troglodytes). Dev. Psychobiol., 34, 119-127.
Bernstein, I. y Sharpe, L. (1966). Social roles in a rhesus monkey group. Behaviour, 26, 91-103. https://doi.org/10.1163/156853966X00038
Black, M. (1970). Margins of precision: essays in logic and language. Ithaca, NY: Cornell University Press.
Boehm, C. (1994). Pacifying interventions at Arnhem Zoo and Gombe. En R. W. Wrangham, W. C. McGrew, F. B. M. de Waal y P. G. Heltne (Eds.), Chimpanzee cultures (pp. 211-226). Cambridge, MA: Harvard University Press.
Bräuer, J., Call, J. y Tomasello, M. (2006). Are apes really inequity averse? Proc. Roy. Soc. Lond. B: Biol. Sci., 273, 3123-3128. https://doi.org/10.1098/rspb.2006.3693
Brosnan, S. F. (2011). A hypothesis of the co-evolution of inequity and cooperation. Front. Decis. Neurosci., 5, 43-55. https://doi.org/10.3389/fnins.2011.00043
Brosnan, S. F. y de Waal, F. B. M. (2003). Monkeys reject unequal pay. Nature, 425, 297-299. https://doi.org/10.1038/nature01963
Brosnan, S. F. y de Waal, F. B. M. (2012). Fairness in animals: where to from here? Soc. Just. Res., 25, 336-351. https://doi.org/10.1007/s11211-012-0165-8
Brosnan, S. F., Schiff, H. C. y de Waal, F. B. M. (2005). Tolerance for inequity may increase with social closeness in chimpanzees. Proc. Roy. Soc. Lond. B: Biol. Sci., 272, 253-258. https://doi.org/10.1098/rspb.2004.2947
Brosnan, S. F., Talbot, C., Ahlgren, M., Lambeth, S. P. y Schapiro, S. J. (2010). Mechanisms underlying responses to inequitable outcomes in chimpanzees, Pan troglodytes. Anim. Behav., 79, 1229-1237. https://doi.org/10.1016/j.anbehav.2010.02.019
Camerer, C. F. y Loewenstein, G. (2004). Behavioral economics: past, present, future. En C. F. Camerer, G. Loewenstein y M. Rabin (Eds.), Advances in behavioral economics (pp. 3-52). Princeton, NJ: Princeton University Press. https://doi.org/0.1515/9781400829118-004
Churchland, P. S. (2011). Braintrust: what neuroscience tells us about morality. Princeton, NJ: Princeton University Press. https://doi.org/10.1515/9781400838080
Coe, C. L. y Rosenblum, L. A. (1984). Male dominance in the bonnet macaque: a malleable relationship. En P. R. Barchas y S. P. Mendoza (Eds.), Social cohesion: essays toward a sociophysiological perspective (pp. 31-63). Westport, CT: Greenwood.
Cords, M. y Thurnheer, S. (1993). Reconciliation with valuable partners by long-tailed macaques. Ethology, 93, 315-325. https://doi.org/10.1111/j.1439-0310.1993.tb01212.x
de Waal, F. B. M. (1987). Tension regulation and nonreproductive functions of sex in captive bonobos (Pan paniscus). Nat. Geogr. Res., 3, 318-335.
de Waal, F. B. M. (1989). Peacemaking among primates. Cambridge, MA: Harvard University Press. https://doi.org/10.4159/9780674033085
de Waal, F.B.M. (1992a). Appeasement, celebration, and food sharing in the two Pan species. En T. Nishida, W. C. McGrew, P. Marler y M. Pickford (Eds.), Topics in Primatology, Vol. 1, Human origins (pp. 37-50). Tokyo: University of Tokyo Press.
de Waal, F. B. M. (1992b). Coalitions as part of reciprocal relations in the Arnhem chimpanzee colony. En A. Harcourt y F. B. M. de Waal (Eds.), Coalitions and alliances in humans and other animals (pp. 233-257). Oxford: Oxford University Press.
de Waal, F. B. M. (1993). Co-development of dominance relations and affiliative bonds in rhesus monkeys. En M. E. Pereira y L. A. Fairbanks (Eds.), Juvenile primates: life history, development, and behavior (pp. 259-270). New York: Oxford University Press.
de Waal, F. B. M. (1996). Good natured: the origins of right and wrong in humans and other animals. Cambridge, MA: Harvard University Press. https://doi.org/10.4159/9780674033177
de Waal, F. B. M. (1998 [orig. 1982]). Chimpanzee politics: power and sex among apes. Baltimore, MD: Johns Hopkins University Press.
de Waal, F. B. M. (2000). Primates — A natural heritage of conflict resolution. Science, 289, 586-590. https://doi.org/10.1126/science.289.5479.586
de Waal, F. B. M. (2006). The tower of morality: reply to commentaries. En S. Macedo y J. Ober (Eds.), Primates & philosophers: how morality evolved (pp. 161-181). Princeton, NJ: Princeton University Press. https://doi.org/10.1515/9781400830336-011
de Waal, F. B. M. (2013). The bonobo and the atheist: in search of humanism among the primates. New York: Norton.
de Waal, F. B. M. y Davis, J. M. (2003). Capuchin cognitive ecology: cooperation based on projected returns. Neuropsychology, 41, 221-228. https://doi.org/10.1016/S0028-3932(02)00152-5
de Waal, F. B. M. y Ren, R. (1988). Comparison of the reconciliation behavior of stumptail and rhesus macaques. Ethology, 78, 129-142. https://doi.org/10.1111/j.1439-0310.1988.tb00224.x
de Waal, F. B. M. y van Roosmalen, A. (1979). Reconciliation and consolation among chimpanzees. Behav. Ecol. Sociobiol., 5, 55-66. https://doi.org/10.1007/BF00302695
Dindo, M. y de Waal, F. B. M. (2006). Partner effects on food consumption in brown capuchin monkeys. Am. J. Primatol., 69, 1-9. https://doi.org/10.1002/ajp.20362
Eberhard, W. G. (1972). The web of Uloborus diversus (Araneae: Uloboridae). J. Zool. Lond., 166, 417-465. https://doi.org/10.1111/j.1469-7998.1972.tb04968.x
Evans, T. A. y Beran, M. J. (2007). Chimpanzees use self-distraction to cope with impulsivity. Biol. Lett., 3, 599-602. https://doi.org/10.1098/rsbl.2007.0399
Fehr, E. y Schmidt, K. M. (1999). A theory of fairness, competition, and cooperation. Q. J. Econ., 114, 817-868. https://doi.org/10.1162/003355399556151
Flack, J. C., Jeannotte, L. A. y de Waal, F. B. M. (2004). Play signaling and the perception of social rules by juvenile chimpanzees. J. Comp. Psychol., 118, 149-159. https://doi.org/10.1037/0735-7036.118.2.149
Flack, J. C., Krakauer, D. C. y de Waal, F. B. M. (2005). Robustness mechanisms in primate societies: a perturbation study. Proc. Roy. Soc. Lond. B: Biol. Sci., 272, 1091-1099. https://doi.org/10.1098/rspb.2004.3019
Gallagher, J. (2004). Evolution? No: A Conversation with Dr. Ben Carson. Adventist Review, 26 (February). https://doi.org/archives/2004-1509/toc.html
Guth, W. (1995). On ultimatum bargaining experiments: a personal review. J. Econ. Behav. Org., 27, 329-344. https://doi.org/10.1016/0167-2681(94)00071-L
Henrich, J., Boyd, R., Bowles, S., Camerer, C., Fehr, E., Gintis, H. y McElreath, R. (2001). In search of Homo economicus: behavioral experiments in 15 small-scale societies. Am. Econ. Rev., 91, 73-78. https://doi.org/10.1257/aer.91.2.73
Herculano-Houzel, S. (2009). The human brain in numbers: a linearly scaled-up primate brain. Front. Hum. Neurosci., 3, 1-11. https://doi.org/10.3389/neuro.09.031.2009
Herrmann, E., Keupp, S., Hare, B., Vaish, A. y Tomasello, M. (2013). Direct and indirect reputation formation in nonhuman great apes (Pan paniscus, Pan troglodytes, Gorilla gorilla, Pongo pygmaeus) and human children (Homo sapiens). J. Comp. Psychol., 127, 63-75. https://doi.org/10.1037/a0028929
Horner, V., Proctor, D., Bonnie, K.E., Whiten, A. y de Waal, F.B.M. (2010). Prestige affects cultural learning in chimpanzees. PLoS-ONE, 5, e10625. https://doi.org/10.1371/journal.pone.0010625
Horowitz, A. (2009). Disambiguating the “guilty look”: salient prompts to a familiar dog behaviour. Behav. Process, 81, 447-452. https://doi.org/10.1016/j.beproc.2009.03.014
Hume, D. (1985 [orig. 1739]). A treatise of human nature. Harmondsworth: Penguin.
Huxley, T.H. (1989 [orig. 1894]). Evolution and ethics. Press, Princeton, NJ: Princeton University.
Jensen, K., Hare, B., Call, J. y Tomasello, M. (2006). What’s in it for me? Self-regard precludes altruism and spite in chimpanzees. Proc. Roy. Soc. Lond. B: Biol. Sci., 273, 1013-1021. https://doi.org/10.1098/rspb.2005.3417
Killen, M. y Rizzo, M. (2014). Morality, intentionality, and intergroup attitudes the origins of morality. Behaviour, 151, 337-359. https://doi.org/10.1163/1568539X-00003132
Kitcher, P. (2006). Ethics and evolution: how to get here from there. En S. Macedo y J. Ober (Eds.), Primates & Philosophers: How Morality Evolved (pp. 120-139). Princeton, NJ: Princeton University Press. https://doi.org/10.1515/9781400830336-009
Kitcher, P. (2014). Is a naturalized ethics possible? Behaviour, 151, 245-260. https://doi.org/10.1163/1568539X-00003145
Koyama, N. F. y Dunbar, R. I. M. (1996). Anticipation of conflict by chimpanzees. Primates, 37, 79-86. https://doi.org/10.1007/BF02382923
Kummer, H. (1995). The quest of the sacred baboon. Princeton, NJ: Princeton University Press.
Logue, A. W. (1988). Research on self-control: an integrating framework. Behav. Brain Sci., 11, 665-709. https://doi.org/10.1017/S0140525X00053978
Mischel, W., Ebbesen, E. B. y Raskoff Zeiss, A. (1972). Cognitive and attentional mechanisms in delay of gratification. J. Person. Soc. Psychol., 21, 204-218. https://doi.org/10.1037/h0032198
Osvath, M. y Osvath, H. (2008). Chimpanzee (Pan troglodytes) and orangutan (Pongo abelii) forethought: self-control and pre-experience in the face of future tool use. Anim. Cogn., 11, 661-674. https://doi.org/10.1007/s10071-008-0157-0
Palagi, E., Paoli, T. y Borgognini Tarli, S. (2006). Short-term benefits of play behavior and conflict prevention in Pan paniscus. Int. J. Primatol., 27, 1257-1270. https://doi.org/10.1007/s10764-006-9071-y
Price, S. A. y Brosnan, S. F. (2012). To each according to his need? Variability in the responses to inequity in non-human primates. Soc. Justice Res., 25, 140-169. https://doi.org/10.1007/s11211-012-0153-z
Proctor, D., Williamson, R. A., de Waal, F. B. M. y Brosnan, S. F. (2013). Chimpanzees play the ultimatum game. Proc. Natl. Acad. Sci. USA, 110, 2070-2075. https://doi.org/10.1073/pnas.1220806110
Range, F., Leitner, K. y Virányi, Z. (2012). The influence of the relationship and motivation on inequity aversion in dogs. Soc. Justice Res., 25, 170-194. https://doi.org/10.1007/s11211-012-0155-x
Roma, P. G., Silberberg, A., Ruggiero, A. M., y Suomi, S. J. (2006). Capuchin monkeys, inequity aversion, and the frustration effect. J. Comp. Psychol., 120, 67-73. https://doi.org/10.1037/0735-7036.120.1.67
Russell, Y. I., Call, J. y Dunbar, R. I. M. (2008). Image scoring in great apes. Behav. Process, 78, 108-111. https://doi.org/10.1016/j.beproc.2007.10.009
Singer, P. (1973). The triviality of the debate over “is-ought” and the definition of “moral”. Am. Philos. Q., 10, 51-56.
Smith, A. (1937 [orig. 1759]). A theory of moral sentiments. New York, NY: Modern Library.
Subiaul, F., Vonk, J., Barth, J. y Okamoto-Barth, S. (2008). Chimpanzees learn the reputation of strangers by observation. Anim. Cogn., 11, 611-623. https://doi.org/10.1007/s10071-008-0151-6
van Hooff, J. A. R. A. M. (1972). A comparative approach to the phylogeny of laughter and smiling. En R. A. Hinde (Ed.), Non-verbal communication (pp. 209-241). Cambridge: Cambridge University Press.
van Wolkenten, M., Brosnan, S. F. y de Waal, F. B. M. (2007). Inequity responses of monkeys modified by effort. Proc. Natl. Acad. Sci. USA, 104, 18854-18859. https://doi.org/10.1073/pnas.0707182104
Verbeek, P., Hartup, W. W. y Collins, W. C. (2000). Conflict management in children and adolescents. En F. Aureli y F. B. M. de Waal (Eds.), Natural conflict resolution (pp. 34-53). Berkeley, CA: University of California Press. https://doi.org/10.1525/9780520924932-006
Vollmer, P. J. (1977). Do mischievous dogs reveal their “guilt”? Vet. Med. Small Anim. Clin., 72, 1002-1005.
von Rohr, C. R., Koski, S. E., Burkart, J. M., Caws, C., Fraser, O. N., Ziltener, A. y van Schaik, C. P. (2012). Impartial third-party interventions in captive chimpanzees: a reflection of community concern. PLoS ONE, 7, e32494. https://doi.org/10.1371/journal.pone.0032494
Westermarck, E. (1917[1908]). The origin and development of the moral ideas. Vol. 2. London: Macmillan.
1. Traducción: E. Joaquín Suárez-Ruíz y Laura Danón. Esta traducción fue autorizada tanto por su autor, Frans de Waal, como por su editorial original, Behaviour (Brill).
2. N. de los T.: Se utilizó la traducción de Félix Duque. Hume, D. 1992 (1739). Tratado De La Naturaleza Humana. Tecnos.
3. N. de los T.: de Waal se refiere al siguiente texto: Kitcher, P. (2014). Is a naturalized ethics possible? Behaviour, 151(2-3), 245-260.